ノンレム睡眠は、眠りの深さに応じて段階がある。
ここで は、3段階(N1・N2・N3)とし、解説します。 【ア】
N1は うとうとの状態で、脳波をみれば、覚醒時のベータ波は消失し、アルファ波を開始します。
※覚醒時でも、安静にしているとアルファ波です。ベータ波より、ゆっくりとした波長です。
筋活動は徐々に低下し、居眠りが始まる。
覚醒時と同じようなベータ波も 生じますが、やがてシータ波へ移行していきます。
眠りが進み、N2に なると、少 しゆっくりとした4~7Hzのシータ波が全般を占めますが、
間隔をおいて、繰り返し、紡錘波やK- 複合波といった、特徴的な脳波が現れます。 【イ】
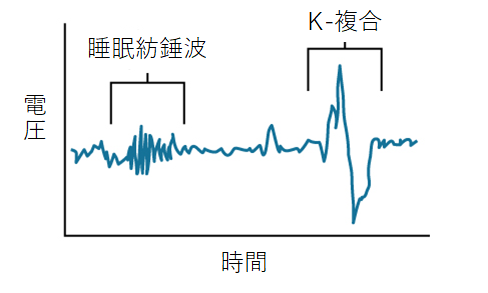
K-複合波は外部刺激に対する反応として、睡眠の安定性を確保するために突発的に発生します。
もっとも深いノンレム睡眠であるN3で は、 さらにゆっくりとした1~3Hzのデルタ波が現れます。
N2と同様に紡錘波(スピンドル波)も発生し、11~16㎐で0.5~3秒間持続します。
スピンドル波は視床と大脳皮質の間で発生する周期的な神経細胞活動の同期によって発生しています。
全般的には大きなデルタ波(徐波)がほとんどの時間を占めるようになり、徐波睡眠ともいいます。 【ウ】
※大きなデルタ波になるのは、点在するニューロンクラスターが同期し、なおかつ共鳴により、振幅が大きくなる からです。
※覚醒時はニューロンがそれぞればらばらに興奮したり、抑制したりしているので、
個々の電気活動が互いに干渉し合い、振動が増したり、振動を打ち消し合ったりしています。
結果として、周期が短く、振幅の小さい脳波(ベータ波)となります。【ウ】
※ガンマ波は、30~100Hzの脳波で、認知機能や注意、記憶に関連し、脳の異なる領域間の情報統合を
円滑に処理し、問題解決や意識の統合を図るなど特に集中しているときに活発になります。 【ウ】
睡眠サイク ルを繰り返すにしたがい、N3が消失する。REMの継続時間は増えてい く。
※N3が果たしている役割(脳の休息)が完了 しているということかもしれない。
また、睡眠の終末においては、REM→N2へ推移するなど、N1の省略もみられる。
これは、記憶へのフラグ付けが完了しており、その作業をする必要がなくなったことを意味しているものと思われる。 【ア】
【ア】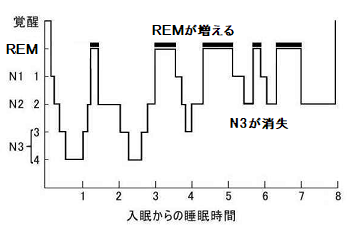 |
【イ】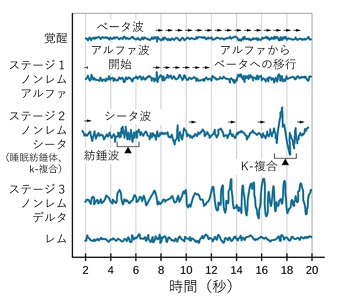 |
スピンドル波 と K-複合
|
【ウ】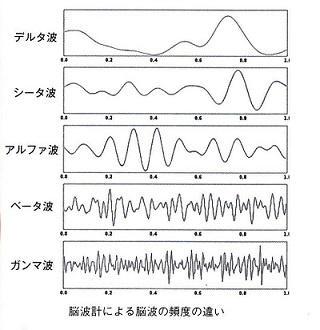 |
ノンレム睡眠中は、筋活動に加え、心拍、呼吸、体温など
自律神経系を介したさまざまな生理活動は低下し安定します。
① これは、身体が休息状態にあることを反映しています。
② デルタ波は、大脳皮質の活動の低下を反映してお り、脳も休息していると言えます。
③ 脳機能の維持や修復、記憶の固定と いった重要な生理的役割も担っています。
睡眠中の脳活動は、睡眠段階によって特徴的なパターンを示しますが、
特にノンレム睡眠において観察される「紡錘波=スピンドル波」と「K複合体」は注目すべきです。
スピンドル波は視床網様核の抑制性ニューロンと視床中継核の興奮性ニューロンが相互に作用し、皮質に投射されることで発生します。
言い換えると、残すべき新奇記憶とそうでない削除すべき短期記憶を選り分け(フラグを立て)ながら、
深層ニューロンの大規模なニューロンの同期活動に関与しているものと思われます。つまり、長期記憶へのアクセスです。
【脳活動と睡眠】 脳のモード
脳のモードと、それをコントロールするサリエンス・ネットワークについて解説しましょう。
SNは 身体の状態を検知し、偏桃体(情動)や腹側被蓋野(報酬系)と連携し、モニタリングします。
意識化により、脳のモード を切り替え、恒常性を維持するための処理も 併行して開始します。 【エ】
ご存じのように、覚醒時で集中しているときはCENモードで す。 ※青い部分 【オ】
覚醒時であっても、ぼんやりしているときはDMNモー ドです。 ※赤い部分 【オ】
CEMとDMNのモード切替をしているのがSNです。 ※黄色い 部分 【オ】
覚醒時は「瞬き」をするように、モードを切り替える様子が確認されています。
睡眠時はSNが 感覚情報(島皮質)を取り込みながら、
偏桃体(感情)と側坐核(報酬系)の連携で、モニタリングし、ネットワークを調整しています。 【エ】
ノンレム睡眠時はSNが主導し、 段階ごとに、脳モードの切り替えを行っています。 【オ】
レム睡眠時は、SNは 背景となり、感情や認識の枠組みをモニタリングしており、DMNが 主活動領域となる。
【エ】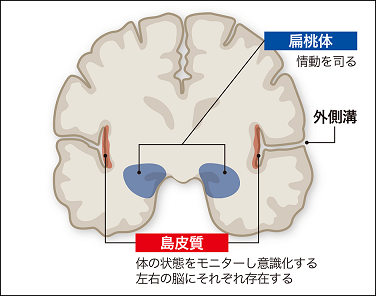 Salience Network では 島皮質に感覚情報を取り込み偏桃体と連携して、 身体の状態に関連付けてモニタリングしている。 |
【オ】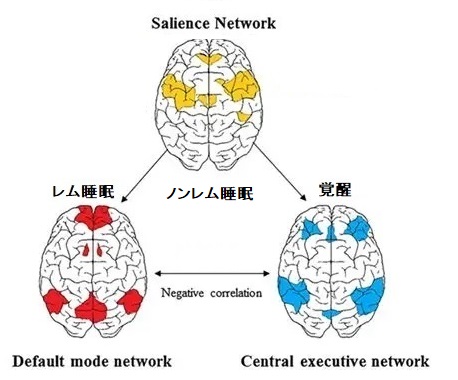 |
①入眠時は、SN状態であり、昼の残滓にかかる蓄積された感覚情報を
島皮質がモニターし、偏桃体が感情を検知します。
②ノンレム1で は、残すべき短期記憶にフラグをたてる際に、CEN⇔DMN状 態となります。※1
アルファ波からベータ波に移行する部分があり、覚 醒に近い活動をしているものと思われる。
③ノンレム2の段階で は、DMNが深化し、 新奇記憶を再生し、長期記憶化へ向けた作業をすすめている。
紡錘波やK -複合といった脳波が生じている。これは新奇記憶へ のアクセスと再生を意味している。※2
④ノンレム3では、不要な短期記憶を消去する際は、DMN状態で、不要なシナプス結合が消滅します。
スピンドル波があるので記憶の定着、全般はデルタ波により不要な記憶の消去が行われている。
デルタ波の振幅を大きくし、 脳内の不要物を除去する作業もしています。※3
⑤ レム睡眠時は、重要な情報を長期保存するため、海馬から「大脳皮質」へと転送します。
この海馬から大脳皮質への記憶の転送作業が、ある程度の期間、繰り返し行わ れています。
⑥ レム睡眠時は脳の血流がノンレム睡眠時と比較して2倍に なります。
海馬におけるニューロンの新生と脳細胞の修復に費やされているものと思われます。
※1 CEN⇔DMN を オートパイロットモード=APM ともいう。
※2 紡錘波(スピンドル波)の最もよく知られた生理的意義は、記憶の固定へ の関与です。
特に宣言的記憶(エピソード記憶や意味記憶など)の固定において重要な役割を果たしています。
覚醒中に獲得された新しい情報は、海馬に一時的に保持されますが、
睡眠中のスピンドル波の発生は、この情報を皮質に転送し、
長期記憶として定着させる プロセスを促進すると考えられています。
K-複合は皮質の深層ニューロンにおける大規模な同期活動に 関与しており、
共鳴により、弱いつながり記憶を 関連付け、ニューロンネットワークの形成に関与していると思われます。
※3 睡眠サ イクルが進むにつれ、N3が消失し、ノンレム睡眠ステップの省略およびREMの増加が見られます。
これは、不要なシナプス結合の解放が完了し、SNが脳のモードを切り替えているものと思われる。